
Moon Illumination & Sasquatch Activity Levels

At the time of publication, over 650 reports have been added to the Sasquatch Data Project dataset. Figures and data used in this blog are from a previous version of the dataset.
Introduction
Fluctuations in moonlight intensity greatly influence the behaviors of both prey and predators (Price, Waser & Bass 1984). The influence of moonlight on sasquatch activity has been a topic of debate among bigfoot researchers for decades, yet there has not been a data-driven study to explore this phenomenon. The Sasquatch Data Project is stepping in to put data at the forefront of this discussion. Part of the mission of the Sasquatch Data Project is to bring clarity to the behavior, appearance, and social dynamics of sasquatches through data from witness testimony. This blog will be looking into the activity levels of sasquatches in relation to moon illumination to explore the possibility of correlation between the two in an effort to learn more about the complex behavioral systems of sasquatches.
Methods
The Sasquatch Data Project is currently building its dataset from the Bigfoot Field Researchers Organization (BFRO) website’s expansive collection of witness reports. The reports, which are in plain text format on the website, are meticulously sifted through to extract as much information as possible, then organized into a spreadsheet optimized for data analysis and coding. At the time of writing, a total 602 reports have been examined, and 16 states that have been documented including Alaska, Alabama, Arkansas (incomplete), Delaware, Maine, Mississippi, Nebraska, Nevada, New Hampshire, North Carolina, North Dakota, Oregon, Rhode Island, South Dakota, Vermont, and Wyoming (incomplete). While moon phase and illumination are not typically stated in the reports, they can be easily obtained through a moon phase “calculator” when an exact date of the encounter is given. Thus, for the majority of reports, that is how the illumination percentage and phase are found. Within the dataset, a classification system has been devised to generally describe the encounter detailed in the witness report. I have identified 5 different “Observation Types” and they are as follows:
Animal: A clear view of a sasquatch was seen by the witness.
Vocalization: A proposed sasquatch vocalization was heard by the witness. I.e. scream, howl, grunt, whistle, etc.
Activity: General sasquatch activity was reported. I.e. bipedal walking/running noises, tree shaking, intimidation displays, rock throwing, etc. A sasquatch was not seen.
Footprint: A footprint(s) was found in substrate by the witness.
Wood Knock: A distinct wood-on-wood sound (or similar) was interpreted by the witness.
Of all the reports, these are the main groups I have identified that are efficient in giving a broad overview of the witness’s experience. These groups are further defined into subcategories called “Encounter Type” (road crossing, camping activity, intimidation display, etc.), but those are not currently relevant for this particular investigation. Due to current data limitations, a more general sense of sasquatch activity will be considered.
Limitations
Before proceeding, it is important to note limitations in the dataset. The type of data the BFRO provides is known as “presence-only” data. This represents only when an event was reported, not every time the event occurred in nature. A person had to experience it, then report it to the BFRO to be recorded. Not everyone does this because they either (1) simply do not want to, (2) do not know such a manner of reporting to an organization exists, or (3) some other completely valid reason. Although it is ideal to have data recorded during non-event periods, the use of presence-only data is entirely acceptable.
Additionally, there is inherent error within witness testimony. To many, having an experience with a sasquatch is extremely distressing, thus affecting their ability to retain particular details and potentially introducing false memories due to the stress and trauma (Kaplan et al. 2015). Witnesses also may not record their experience immediately following, introducing inaccuracies in details and dates. While witness testimony is not completely concrete, the bulk of current sasquatch data lies within it, therefore we must acknowledge these inaccuracies and proceed in investigating.
Data
At the time of writing, 602 reports from the BFRO dataset have been parsed. Of these 602 reports, 272 reports described by the 5 main “Observation Types” have included an exact date therefore allowing a moon illumination percentage to be obtained. Also, for the sake of clarity, when referencing the “Activity” category from the dataset, I will always capitalize the word. When referencing general sasquatch activity (this could include a vocalization, sighting, or any general encounter type), it will not be capitalized. I recognize this can be confusing and will rename the category at a later date.

Figure 1. Counts of observation occurrences for the main observation types and their corresponding moon illumination percentage values. Data is not filtered for time.
Figure 1 includes all 272 reports, showcasing the counts of specific Observation Types and their corresponding moon illumination percentage grouped into 20% intervals. Grouping moon illumination values in either 20% or 25% intervals is typical and a valid way to bin the data. This figure does not take into account the time of day and shows the overall trends regardless of time. Data filtered by time will be introduced in Figures 2 & 3. It is also important to note Google Sheets does not incorporate the upper bound of the group into the bin. The group 0-20% includes moon illumination values of 0-19.9%. The group 20-40% includes percentages 20-39.9%, and so on. The groups start at the first value and go up to, but do not include, the upper bound. The only exception to this is the last group 80-100% where both values are included in the bin.
Illustrated in Figure 1, the data is prominently U-shaped, showing peaks in frequencies of all observation types under the extreme low (0-20%) and high (80-100%) moon illumination conditions, and substantially lower occurrence counts within the intermediate groupings. Most notably, the number of sasquatch sightings and vocalization reports peak in both the high and low ranges, exceeding double the number of occurrences compared to the intermediate moon illumination groups. The counts of Observation Types for all categories, except “Activity”, reaches a minimum in the 40-60% range. Isolated wood knocks are completely absent from this moon illumination range.
Figure 1B confirms we have U-shaped data through the very high R^2 values of the polynomial trendlines. R^2 values represent how well the equation of the trendline fits the data. The closer to 1 the better. In our case, R^2 values greater than 0.9 indicate less than a 10% chance the model cannot explain the variance in the data, meaning the data it can predict is not likely up to random chance. R^2 values for the “Animal”, “Vocalization”, and “Activity” types come in at 0.943, 0.954, and 0.901, respectively. Th R^2 values could potentially be significant if the values hold strong as more data is added to the dataset. Because of the current lack of data regarding the “Footprint” and “Wood Knock” Observation Types, these values will be disregarded for the remainder of the blog.

Figure 2. Counts of Main Observation Types and their corresponding moon illumination percentages for times 8:00PM to 6:00AM.
Figure 2 depicts the 129 data points of nighttime occurrences of each primary observation type along with its corresponding moon illumination percentage. For this study, nighttime is defined as 8:00PM to 6:00AM, but not including 6:00AM. This timeframe was selected in an effort to only look at times when the moon’s light would be most prominent. It is important to note these times change throughout the year with earlier and later sunrises and sunsets. Until more data is available, this is the chosen method of filtering the data.
The same general trends regardless of time of day follow into the night data. The main differences being the stark decrease in sasquatch sightings in the 0-20% group compared to the other categories. The majority of sasquatch sightings and general activity are happening under high moon illumination values in the 80-100% range. The majority of vocalizations are held within the 0-20% and 80-100% ranges.
The trendlines again see strong R^2 values, this time coming in at 0.91, 0.966, and 0.801 for the “Animal”, “Vocalization”, “Activity” categories, respectively. The “Activity” observation type dropped by 0.1, but in the biological sciences an R^2 value above 0.7 is generally still considered strong, and even 0.5 can be acceptable. Again, the model is efficient in fitting the data.
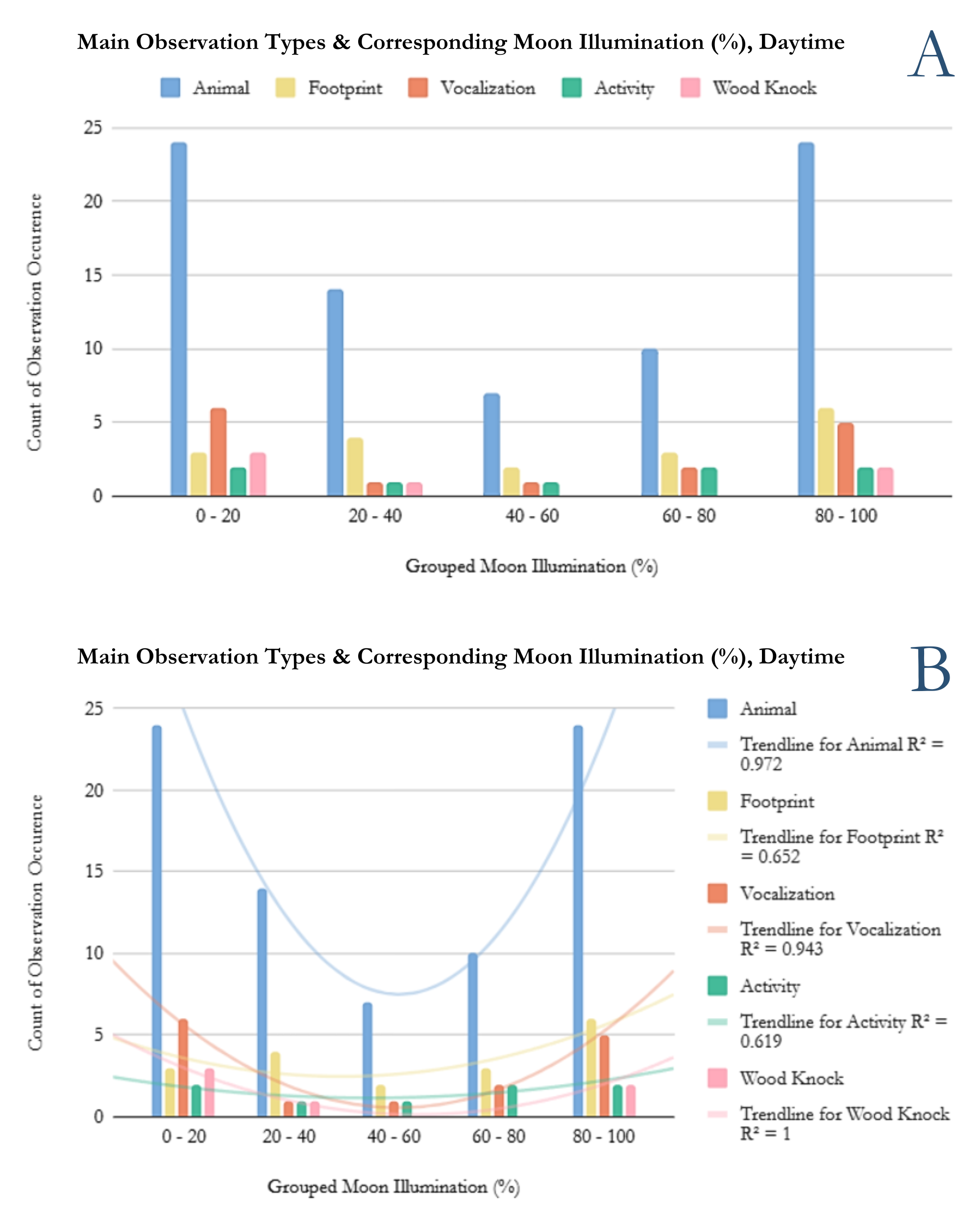
Figure 3. Counts of Main Observation Types and their corresponding moon illumination percentages for times 6:00AM to 8:00PM.
Figure 3 includes the 125 reports of observation occurrences with moon illumination values during the daytime. The daytime timeframe is defined as 6:00AM to 8:00 PM. In this case, reports explicitly stating 8:00PM and beyond are not included. Figure 3 greatly follows the general trend of Figure 1 for the animal sightings category. The biggest differences between Figure 1 and 3 being the stark decrease in vocalizations for both the 0-20% and 80-100% groups.
For this subset of data, R^2 values are 0.972, 0.943, and 0.619 for the “Animal”, “Vocalization”, and “Activity” observation types, respectively. While the “Animal” and “Vocalization” categories remain very strong, “Activity” waivers. The model does not describe the “Activity” data as well, yet could still be regarded as acceptable as more data is collected.

Figure 4. A bar chart representing the different eye shine colors reported in visual sightings from times 8:00PM to 6:00AM.
Figure 4 depicts the range of eye shine colors, if present at all, from the 19 visual sighting reports that occurred between 8PM and 6AM and stated the tapetal reflex (eye shine) color. Of these 19 sightings, 14 reports mention noticing eye shine, while 5 reported its absence. Eye shine colors that have been reported are red, yellow, white, green, yellow-green, orange, and gold. It could be argued that yellow and gold could be combined into the same group, as well as green and yellow-green, but for now they will remain separate categories.
Discussion
There is a clear correlation between moon illumination and sasquatch activity with the current dataset. The preliminary results to be discussed should be taken as just that, preliminary. The point of this blog is to show what the data currently suggest, and to learn how models change as more data becomes available. That said, there is sufficient data to begin looking into potential trends and identifying points of interest within sasquatch behavior.
It is obvious from the data collected thus far that sasquatch encounters are commonly occurring during low (0-20%) and high (80-100%) moon illumination regardless of time of day. This in itself is particularly interesting in that even when the moon is not the prominent light source (i.e. daytime) its influence holds strong during these phases. Evaluating why sasquatches seem more active during the extremes is going to be purely based on speculation and the habits of scientifically recognized nocturnal predators.
Beginning with the nighttime data, all three main Observation Type categories, “Animal”, “Vocalization”, and “Activity”, reached their maximums under high moon illumination (80-100%). The increase in sasquatch sightings under these conditions can probably be attributed to a few factors. First and foremost, humans can see the best under high moon illumination. Ultimately, a human must observe and then report a sasquatch to “officially” have a sighting. Consequently, more reports would come in under these conditions since humans lack night vision and rely on the moon’s light at night to aid our vision.
The specific reasons as to why sasquatches would be more active during times of high moon illumination require a look into known predator-prey relationships. Nocturnal predators tend to use the increased visibility of high moon illumination nights to both patrol their territory and hunt. Prey rely on both direct (predator sightings, scat, moonlight, etc.) and indirect cues to assess predation risk, significantly influencing their nocturnal activity levels (Nersesian, Banks, & McArthur 2011). Increased predation risk during nights with higher moon illumination has caused some species of prey to reduce activities by 40-70% in an effort to increase their chances of survival (Taylor et al., 2023). Predator-prey relationships are extremely complex, and details vary among species, but the general decrease in activity remains true for many prey mammals. Nocturnal predators are also known to patrol their territory during periods of high moon illumination as the decrease in prey activity makes hunting a more energy taxing activity. Coyotes, gray wolves, and mountain lions are just a few of the large nocturnal predators in North America that exhibit this behavior. Nocturnal predators are also known to exhibit more frequent vocalizations to aid in claiming territory and signal to others of their kind during a hunt. Figure 2 details an increase in reported sasquatch vocalizations during both high and low moonlight nights, tentatively supporting this idea. With the current data at hand, the types of encounters in conjunction with high moon illumination can currently be explained by known predator behavior.
Under low (0-20%) moon illumination conditions, sightings of sasquatches decrease compared to the high moon illumination reports but are still prominent compared to the entirety of the dataset. This is potentially partly due to a human’s inability to see as well in darker conditions. If a human cannot see, a sighting cannot happen even if the animal is in close proximity. The decrease in sightings does not necessarily mean sasquatches are not as active with low moon illumination as seen with the numerous reports of vocalizations during these periods. Low light could cause sasquatches to rely more on their voices to communicate with one another, or could be used to signal different stages of a hunt as seen in chimpanzees (Mine et al. 2022). Low light conditions make for the perfect hunting conditions, due to prey perceiving less of a predation risk under these conditions (Taylor et al. 2023). Great apes also vocalize while hunting to ward off other predators in the area and intimidate prey. Various factors, including those not mentioned, could contribute to the increase in vocalizations from sasquatches during periods of low moon illumination.
Even when moon illumination would not influence visual quality (i.e. the daytime), sasquatch activity continues to follow the same general trend of peaks during times of high and low moonlight (Figure 3). This could potentially point to sasquatches having a more dynamic and flexible circadian rhythm over a simple day-night pattern. This could also be an adaptation to the lunar cycles of certain prey species and to avoid competition with other predators like black bears, mountain lions, and coyotes. Additionally, sasquatches are reported to be massive animals. They likely require an enormous caloric intake each day to sustain themselves. For example, a typical gorilla consumes approximately 4,000 calories a day. Orangutans eat around 3,000 calories a day, but will consume upwards of 8,500 when certain foods, like fruit, are plentiful (Knott 1997). Therefore, it is not outlandish to assume sasquatches would also require a similar caloric intake, and probably more, due to their massive, muscular size. Potentially, this alone could make it necessary for sasquatches to hunt day and night. If these preliminary findings hold true as more data is parsed, is it even fair to call sasquatch a nocturnal animal? Currently it is too early to say conclusively, but it is an intriguing question to pose. If this trend of daytime activity increases during low and high moon illumination periods continues, this could point to sasquatches as having highly complex and flexible behavioral tendencies that do not fit into the traditional mold of diurnal or nocturnal.
A shift in perception of the potential circadian rhythm of sasquatches could be biologically plausible. Great apes are not considered nocturnal animals, though approximately 69% of mammals are (Bennie et al. 2014). Particular chimpanzee groups have been noted as adopting nocturnal tendencies to adapt better to their environment (Tagg et al. 2018), but there are no true nocturnal great apes currently recognized by science. Because of the diurnal nature of great apes, they also lack the presence of a tapetum lucidum, a layer of tissue on the eye that aids sight in low light conditions. The tapetum lucidum is made apparent to the observer by the presence of “eye shine” from the animal. Eye shine appears as an array of colors dependent on the host species. These variations can be caused by different compositions, blood vessels, age, structure, and light source (Ollivier 2004), but individuals within the same species typically have the same color tapetum reflex (eye shine). As stated earlier, sasquatches do not appear to be strictly nocturnal animals, though this does not explain why a number of witnesses have noted an apparent presence of eye shine in certain sighting situations. Eye shine color distributions reported by sasquatch witnesses and their counts can be seen in Figure 4. It is important to note there are witnesses who did not notice eye shine when they theoretically should have, such as seeing a sasquatch’s eyes in their headlights. If sasquatches are adapting their habits to fit their environments and are not necessarily following strict diurnal or nocturnal behaviors, it is not unreasonable to think potentially, and I say this with extreme caution, that some genetically similar groups in particular areas have evolved to have eye shine, while others did not to fit their immediate environmental niches. While in theory this could happen, the evolution of the tapetum lucidum is quite complex and involves multiple genes to be present. For vertebrates, there are two main types of the tapetum lucidum, with mammals exclusively having the choroidal tapetum. The different types of tapetum lucidum evolved independently of one another yet their structure and function are remarkably similar (Schwab et al. 2002). That said, no great apes possess a tapetum lucidum, and the necessary genes were probably lost in a previous common ancestor. Though lost for most primates, the tapetal reflex did re-evolve in certain nocturnal primates like tarsiers. It is possible sasquatches could have done the same, and is an example of convergent evolution (Schwab et al. 2002). Reports noting an apparent lack of a tapetal reflex could also be explained by the idea that the species as a whole does indeed possess tapetum lucidum, but certain groups or individuals may not have as intense eye shine due to a number of potential factors including viewing angle of the light, stress, age, or any other number of reasons.
Conclusion
The current data suggests a close correlation between moon illumination and sasquatch activity. Whether these trends are due to humanistic habits or the potentially complex circadian rhythms of sasquatches, it is unclear for the time being until more data is parsed.
If you have any questions or nice comments, please feel free to leave them here! You can also reach me by email or the Contact form on this website.